Simone Cauzzo1,2, Kavita Singh2, Matthew Stauder2, Maria Guadalupe Garcia Gomar2, Nicola Vanello3, Claudio Passino1,4, Jeffrey Staab5,6, Iole Indovina7,8, and Marta Bianciardi2
1Life Sciences Institute, Sant'Anna School of Advanced Studies, Pisa, Italy, 2Brainstem Imaging Laboratory, Department of Radiology, Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts General Hospital and Harvard Medical School, Boston, MA, United States, 3Dipartimento di Ingegneria dell’Informazione, University of Pisa, Pisa, Italy, 4Fondazione Toscana Gabriele Monasterio, Pisa, Italy, 5Department of Psychiatry and Psychology, Mayo Clinic, Rochester, MN, United States, 6Department of Otorhinolaryngology – Head and Neck Surgery, Mayo Clinic, Rochester, MN, United States, 7Department of Biomedical and Dental Sciences and Morphofunctional Imaging, University of Messina, Messina, Italy, 8Laboratory of Neuromotor Physiology, IRCCS Santa Lucia Foundation, Roma, Italy
1Life Sciences Institute, Sant'Anna School of Advanced Studies, Pisa, Italy, 2Brainstem Imaging Laboratory, Department of Radiology, Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts General Hospital and Harvard Medical School, Boston, MA, United States, 3Dipartimento di Ingegneria dell’Informazione, University of Pisa, Pisa, Italy, 4Fondazione Toscana Gabriele Monasterio, Pisa, Italy, 5Department of Psychiatry and Psychology, Mayo Clinic, Rochester, MN, United States, 6Department of Otorhinolaryngology – Head and Neck Surgery, Mayo Clinic, Rochester, MN, United States, 7Department of Biomedical and Dental Sciences and Morphofunctional Imaging, University of Messina, Messina, Italy, 8Laboratory of Neuromotor Physiology, IRCCS Santa Lucia Foundation, Roma, Italy
Using high spatial
resolution 7 Tesla resting-state fMRI and a recently developed in-vivo brainstem nuclei atlas, we
report the functional connectome of autonomic and sensory brainstem nuclei in
living humans.
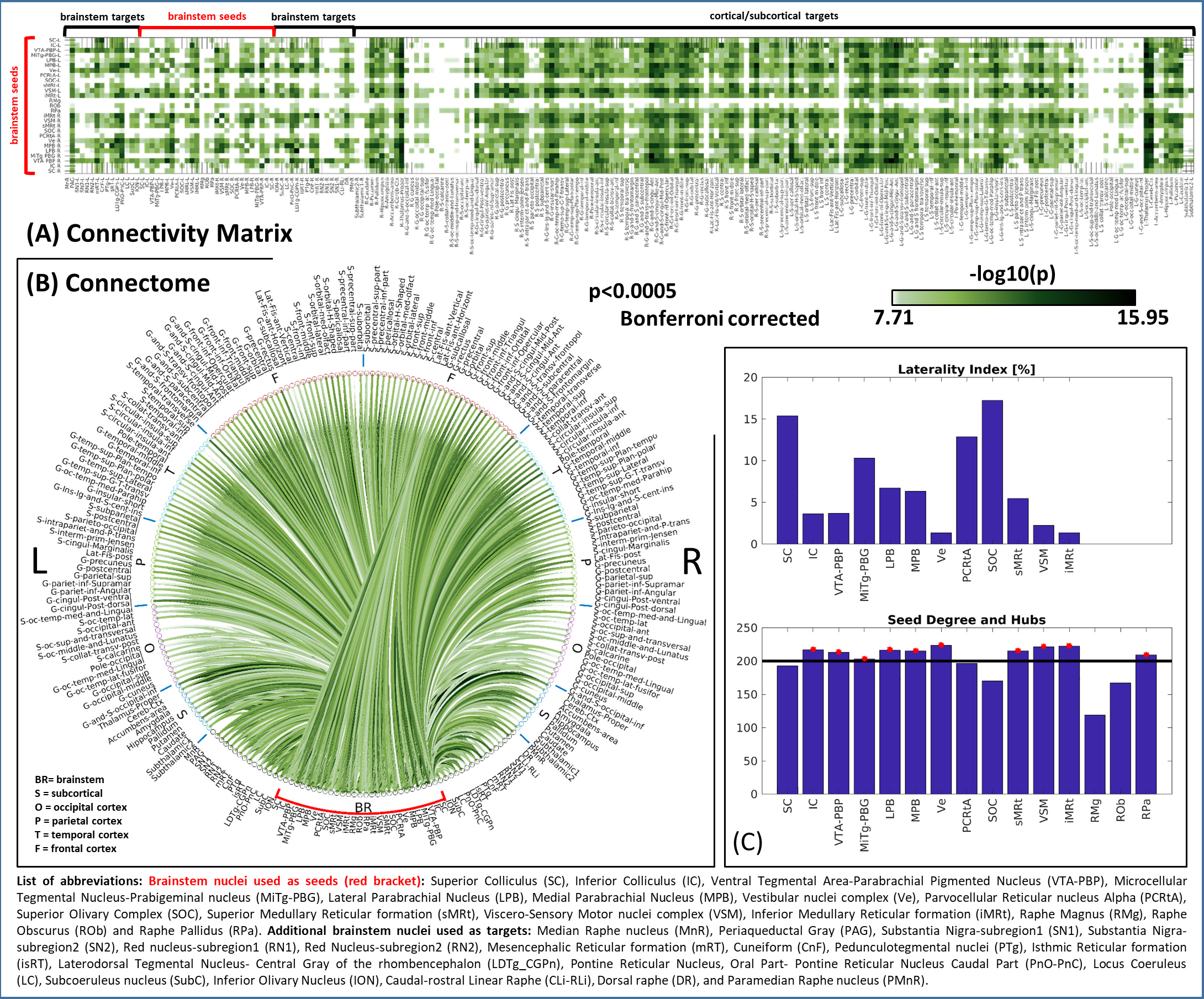
Figure 2: (A) Connectivity
matrix (i.e. mean connectivity values, n = 20) and (B) circular connectome (–log10(p-value))
of 15 brainstem nuclei (red bracket).
For both (A) and (B), for display purposes, we thresholded the results at
p<0.0005 Bonferroni-corrected. C) On top, bar plot displaying laterality
index for bilateral seeds only. On bottom, bar plot displaying the connectivity
degree of each seed: the horizontal line refers to the average degree. Seeds
exceeding the average (i.e. IC, VTA-PBP, LPB, MPB, Ve, sMRt, VSM, iMRT) were considered
hubs and marked with a red star.

Figure 3: Functional connectome of (A) left
iMRt and (B) left MPB (p<0.0005 Bonferroni
corrected for display purposes). (A)
We confirmed iMRt connectivity to: ION, previously found in cats12; Ve13; VSM14 for taste-elicited ingestion/rejection responses;
PAG for pain modulation15. Expected connectivity with SC and LPB were low11. (B) MPB showed expected connectivity to DR, thalamus,
and (weaker) VSM and amygdala, yet we did not observe connectivity to RMg.
Connectivity to insular and cingulate cortex was strong11. Connectivity to PAG and VTA was in line with rat
studies16,17.